- The main growth and development phase in plants begins with the germination of the mature seed.
- Seeds are of two kinds depending on the number of cotyledons or embryo leaves.
Parts of a Seed
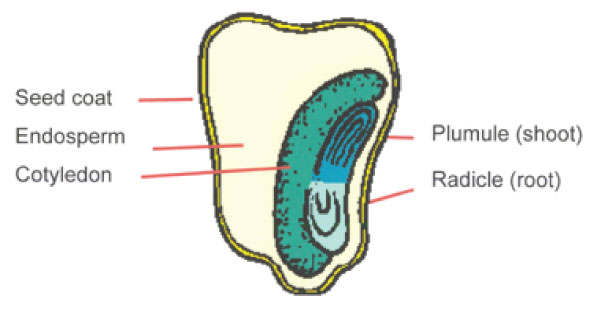
- A typical seed consists of a seed coat enclosing an embryo.
- The seed coat is the outer covering which, in most seeds, is made up of the two layers; an outer testa and inner one, the legmen.
- The testa is thick; the legmen is a transparent membrane tissue.
- The two layers protect the seed bacteria, fungi and other organisms which may damage it.
- There is a scar called hilum, one part of the seed.
- This is point where the seed had been attached the seed stalk or funicle.
- Near one end of 1 hilum is a tiny pore, the micropyle.
- This allows water and air into the embryo, embryo is made up of one or two seed leaves, cotyledons, a plumule (embryonic) and a radicle (the embryonic root).
- The of the radicle is opposite the micropyle.
- In some seeds the cotyledons swollen as they contain stored food for growing plumule and radicle. Such seeds, called non-endospermic seeds.
- In other cases, the seeds have their food stored in: endosperm.
- Such seeds are call endospermic seeds. Seeds with one cotyk are referred to as monocotyledonous while those with two are referred to dicotyledonous.
- This is the major basis differentiation between monocotyledonae and dicotyledonae.
Diagram of a Seed
Maize Grain
Bean Seed
Dormancy in Seeds
- The embryo of a dry, fully developed seed usually passes through a period of rest after ripening period.
- During this time the seed performs all its life (physiological) processes very slowly and uses up little food. This is a period of dormancy.
- Even if all the favourable environmental conditions for germination are provided to the seed during this period of dormancy, the seed will not germinate.
- This is due to the fact that the seed embryo may need to undergo further development before germination.
- Some seeds can germinate immediately after being shed from the parent plant (e.g. most tropical plants) while others must pass through dormancy period, lasting for weeks, months or even years before the seed can germinate.
- Dormancy provides the seeds with enough time for dispersal so that they can germinate in a suitable environment.
- It also enables seeds to survive during adverse environmental conditions without depleting their food reserves.
- The embryo has time to develop until favourable conditions are available e.g. availability of water.
- Embryo may not yet be fully developed.
- Presence of chemical inhibitors that inhibit germination in seeds e.g.abscisic acid.
- Very low concentrations of hormones e.g. gibberellins and enzymes reduces the ability of seeds to germinate.
- Hard and impermeable seed coats prevent entry of air and water in some seeds e.g. wattle.
- In some seeds the absence of certain wavelengths of light make them remain dormant e.g. in some lettuce plants.
- Freezing of seeds during winter lowers their enzymatic activities rendering them dormant.
- When the seed embryos are mature then the seed embryos can break dormancy and germinate.
- Increase in concentration of hormones e.g. cytokinins and gibberellins stimulate germination.
- Favourable environmental factors such as water, oxygen and suitable temperature.
- Some wavelengths of light trigger the production of hormones like gibberellins leading to breaking of dormancy.
- Scarification i.e. weakening of the testa is needed before seeds with hard impermeable seed coats can germinate.
- This is achieved naturally by saprophytic bacteria and fungi or by passing through the gut of animals.
- In agriculture the seeds of some plants are weakened by boiling, roasting and cracking e.g. wattle.
Seed Germination
- The process by which the seed develops into a seedling is known as germination.
- It refers to all the changes that take place when a seed becomes a seedling.
- At the beginning of germination water is absorbed into the seed through the micropyle in a process known as imbibition and causes the seed to swell.
- The cells of the cotyledons become turgid and active.
- They begin to make use of the water to dissolve and break down the food substances stored in the cotyledons.
- The soluble food is transported to the growing plumule and radicle.
- The plumule grows into a shoot while the radicle grows into a root.
- The radical emerges from the seed through micropyle, bursting the seed coat as it does so.
Conditions Necessary for Germination
- Seeds can easily be destroyed by unfavourable conditions such as excessive heat, cold or animals.
- Seeds need certain conditions to germinate and grow.
- Some of these conditions are external, for example water, oxygen and suitable temperature while others are internal such as enzymes, hormones and viability of the seeds themselves.
1. Water
- A non-germinating seed contains very little water.
- Without water a seed cannot germinate.
- Water activates the enzymes and provides the medium for enzymes to act and break down the stored food into soluble form.
- Water hydrolyses and dissolves the food materials and is also the medium of transport of dissolved food substances through the various cells to the growing region of the radical and plumule.
- Besides, water softens the seed coat which can subsequently burst and facilitate the emergence of the radicle.
2. Oxygen
- Germinating seeds require energy for cell division and growth.
- This energy is obtained from the oxidation of food substances stored in the seed through respiration thus making oxygen an important factor in seed germination.
- Seed in water logged soil or seed buried deep into the soil will not germinate due to lack of oxygen.
3. Temperature
- Most seeds require suitable temperature before they can germinate.
- Seeds will not germinate below 0°C or above 47° C.
- The optimum temperature for seeds to germinate is 30°C.
- At higher temperature the protoplasm is killed and the enzymes in the seed are denatured.
- At very low temperatures the enzymes become inactive.
- Therefore, the protoplasm and the enzymes work best within the optimum temperature range.
- The rate of germination increases with temperature until it reaches an optimum.
- This varies from plant to plant.
4. Enzymes
- Enzymes play a vital role during germination in the breakdown and subsequent oxidation of food.
- Food is stored in seeds in form of carbohydrates, fats and proteins which are in insoluble form.
- The insoluble food is converted into a soluble form by the enzymes.
- Carbohydrates are broken down into glucose by the diastase enzyme, fats into fatty acids and glycerol by lipase, and proteins into amino acids by protease.
- Enzymes are also necessary for the conversion of hydrolysed products to new plant tissues.
5. Hormones
- Several hormones play a vital role in germination since they act as growth stimulators.
- These include gibberellins and cytokinins.
- These hormones also counteract the effect of germination inhibitors.
6. Viability
- Only seeds whose embryos are alive and healthy will be able to germinate and grow.
- Seeds stored for long periods usually lose their viability due to depletion of their food reserves and destruction of their embryo by pests and diseases.
Types of Germination
- The nature of germination varies in different seeds.
- During germination the cotyledons may be brought above the soil surface.
- This type of germination is called epigeal germination.
- If during germination the cotyledons remain underground the type of germination is known as hypogeal.
1. Epigeal Germination
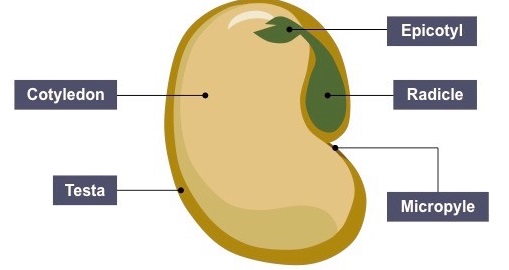
- During the germination of a bean seed, the radicle grows out through the micropyle.
- It grows downwards into the soil as a primary root from which other roots arise.
- The part of the embryo between the cotyledon and the radicle is called the hypocotyl.
- This part curves and pushes upwards through the soil protecting the delicate shoot tip.
- The hypocotyls then straightens and elongates carrying with it the two cotyledons which turn green and leafy.
- They start manufacturing food for the growing seedling.
- The plumule which is lying between two cotyledons, begins to grow into first foliage leaves which start manufacturing food.
2. Hyopgeal Germination
- In maize, the endosperm provides food to the embryo which begins to grow.
- The radicle along with a protective covering grows out of the seed.
- The epicotyl is the part of the embryo between the cotyledon and the plumule.
- The seedling now begins to produce its own food and the endosperm soon shrivels.
- This type of germination in which the cotyledon remains below the ground is known as hypogeal germination.
Primary and Secondary Growth
- The region of growth in plants is found in localised areas called meristems as shown.
- A meristem is a group of undifferentiated cells in plants which are capable of continuous mitotic cell division.
- The main meristems in flowering plants are found at the tips of shoots and roots, in young leaves, at the bases of the inter-nodes, and in vascular cambium and cork cambium.
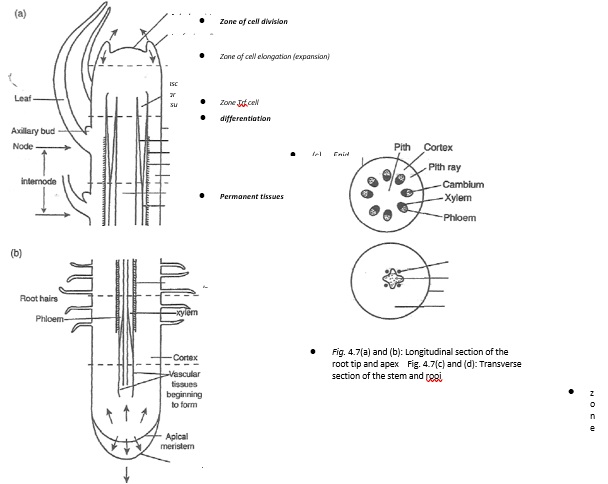
- The meristems at the tips of the shoots and the roots are known as apical meristems and are responsible for primary growth.
- The cambium meristems are responsible for secondary growth.
Primary Growth in Plants
- Primary growth occurs at the tips of roots and shoots due to the activity of apical meristems.
- These meristems originate from the embryonic tissues. In this growth there are three distinctive regions, the region of cell division, cell ejpngarion and cell differentiation.
- The regipn of cell division is an area of actively dividing meristematic cells.
- These cells have thin cell walls, dense cytoplasm and no vacuoles.
- In the region of cell elongation, the cells become enlarged to their maximum size by the stretching of their walls.
- Vacuoles start forming and enlarging. In the region of ceH differentiation the cells attain their permanent size, have large vacuoles and thickened watt cells.
- The cells also differentiate into tissues specialised for specific functions.
- Primary growth results into an increase in the length of shoots and roots.
Secondary Growth in Plants
- Secondary growth results in an increase in width or girth due to activity of the cambium. In secondary growth new tissues are formed by vascular cambium and cork cambium. In monocotyledons plants there are no cambium cell in the vascular bundles. The growth in diameter is due to the enlargement of the primary cells.
- Secondary growth in dicotyledonous plants begins with the division of vascular cambium to produce new cambium cells between the vascular bundles. This forms a continuous cambium ring. These cambium cells divide to form the new cells that are added to the older ones. The cambium cells have now become meristematic.
- The new cells produced to the outer side of cambium differentiate to become secondary phloem and those to the inner side differentiate to become the secondary xylem. More secondary xylem is formed than secondary phloem.
- As a result of the increase in the volume of the secondary tissues, pressure is exerted on the outer cells of the stem. This results in stretching and rupturing of the epidermal cells. In order to replace the protective outer layer of the stem, a new band of cambium cells are formed in the cortex. These cells, called cork cambium orphellogen originate from the cortical cells. The cork cambium divides to produce new cells on either side.
- The cells on the inner side of the cork cambium differentiate into secondary cortex and those produced on the outer side become cork cells. Cork cells are dead with thickened walls. Their walls become coated with a waterproof substance called suberin. The cork cells increase in number and become the bark of the stem. This prevents loss of water, infection from fungi and damage from insects. The corky bark is also resistant to fire and thus acts as an insulatory layer.
- The bark is normally impermeable to water and respiratory gases. Periodically the cork cells, instead of being tightly packed, they form a loose mass. This mass is known as Jenticel.
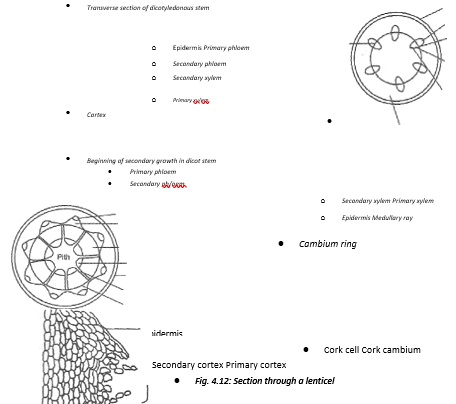
- The rate of secondary growth in a stem varies with seasonal changes.
- During rainy season, xylem vessels and tracheids are formed In large numbers.
- These cells are large, have thin walls and the wood has a light texture.
- In the dry season, the xylem and trancheids formed are few in number.
- They are small, thick-walled and their wood has a dark texture.
- This leads to the development of two distinctive layers within the secondary xylem formed in a year, called annual rings.
- It is possible to determine the age of a tree by counting the number of annual rings.
- Furthermore climatic changes of the past years can be inferred from the size of the ring.
Plants Growth Hormones
- Plant hormones are chemicals produced in very small amounts within the plant body, and play a very important part in regulating plant growth and development. Most growth hormones are produced at the tip of a shoot and transported downwards to the root. The root tip produces very small quantities of the hormones.
- There are many different types of plant hormones and one well-known group is the auxins. Indoie acetic acid (IAA) is one best known auxin. Auxins are produced at the shoot and root tips. Maximum influence on growth in plants occurs when auxins are produced simultaneously with other plant hormones e.g. gibberellins. Maximum growth response in stems requires more IAA than in roots.
- Auxins are known to have various effects on the growth and development in plants. They stimulate cell division and cell elongation in stems and roots leading to primary growth. Auxins cause tropic responses, which are growth responses in plants due to external stimuli acting from a given direction.
- On the other hand IAA stimulates the growth of adventitious roots which develop from the stem rather than the main root. Cuttings can be encouraged to develop roots with the help of IAA. If the cut end of a stem is dipped into IAA, root sprouting is faster. IAA is also used to induce parthenocarpy. This is the growth of an ovary into a fruit without fertilisation. This is commonly used by horticulturalists to bring about a good crop of fruits particularly pineapples.
- Auxins are known to inhibit development of side branches from lateral buds. They therefore enhance apical dominance. During secondary growth auxins Play an important role by initiating cell division in the cambium and differentiation of these cambium cells into vascular tissues.
- Auxins in association with other plant hormones such as the cytokinins induce the formation of callus tissue which causes the healing of wounds. When the concentration of auxins falls in the plant, it promotes formation of an abscission layer leading to leaf fall.
- Gibberellins are another important group of plant growth hormone. Gibberellins are a mixture of compounds and have a very high effect on growth. The most important in growth is gibberellic acid. Gibberellins are distinguished from auxins by their stimulation of rapid cell division and cell elongation in dwarf varieties of certain plants. Dwarf conditions are thought to be caused by a shortage of gibberellins due to a genetic deficiency.
- Gibberellins are important in fruit formation. They induce the growth of ovaries into fruits after fertilisation. They also induce parthenocarpy. Gibberellins also promote formation of side branches from lateral buds and breaks dormancy in buds. This is common in species of temperate plants whose buds become dormant in winter. In addition, this hormone also inhibits sprouting of adventitious roots from stem cuttings, it retards formation of abscission layer hence reduces leaf fall. Gibberellins also break seed dormancy by activating the enzymes involved in the breakdown of food substances during germination.
- Cytoknins also known as kinetins, are growth substances which promote growth in plants when they interact with auxins. In the presence of auxins, they stimulate cell division thereby bringing about growth of roots, leaves and buds. They also stimulate formation of the callus tissues in plants.
- The callus tissue is used in the repair of wounds in damaged parts of plants.
- Cytokinins promote flowering and breaking of seed dormancy in some plant species. They also promote formation of adventitious roots from stems and stimulate lateral bud development in shoots. When in high concentration cytokinins induce cell enlargement of leaves but in low concentration they encourage leaf senescence and hence leaf fall.
- Ethylene is a growth substance produced in plants in gaseous form. Its major effect in plants is that it causes ripening and falling of fruits. This is widely applied in horticultural farms in ripening and harvesting of fruits. It stimulates formation of abscission layer leading to leaf fall, induces thickening of stems by promoting cell division and differentiation at the cambium meristem. But it inhibits stem elongation. Ethylene promotes breaking of seed dormancy in some seeds and flower formation mostly in pineapples.
- Abscisic acid is a plant hormone whose effects are inhibitory in nature. It inhibits seed germination leading to seed dormancy, inhibits sprouting of buds from stems and retards stem elongation. In high concentration, abscisic acid causes closing of the stomata. This effect is important in that it enables plants to reduce water loss. It also promotes leaf and fruit fall. Another hormone, florigen is produced in plants where it promotes flowering.
Apical Dominance
- Although auxins, particularly IAA are important stem and root elongation, they are known to exert profound effects on other aspects of plant growth and development.
- If an apical bud which normally contains high concentrations of auxins is removed, it is observed that more lateral buds lower down the stem sprout, producing many branches.
- This shows that high concentrations of auxins have an inhibitor effect on sprouting of lateral buds and therefore hinders growth of many branches.
- This forms the basis of pruning in agriculture where more branches are required for increased harvest particularly on crops like coffee and tea.
- The failure of lateral buds to develop in the presence of an apical bud is due to the diffusion of auxins from the shoot apex downwards in concentrations higher than that promoting lateral bud development.